1. Introduction
Lipid droplets (LDs) were once viewed as inert blobs of stored fat—simple depots of excess calories kept on standby for future energy needs. However, modern cellular biology has radically transformed this outdated view. Today, lipid droplets are recognized as dynamic, metabolically active organelles central to energy homeostasis, hormonal communication, inflammatory signaling, and metabolic flexibility. They are essential not only for energy storage but also for coordinating how cells adapt to nutrient excess, nutrient scarcity, oxidative stress, immune activation, and endocrine cues.
LD biology sits at the intersection of metabolism, endocrinology, structural cell biology, and nutrition science. Understanding how lipid droplets grow, shrink, communicate with mitochondria, and regulate lipid trafficking offers a powerful framework for understanding obesity, insulin resistance, muscle metabolism, liver health, and even aging. Far from being passive storage spheres, LDs act as miniature biochemical command centers that dynamically expand, contract, fuse, relocate, and modulate cellular function in response to physiological needs.
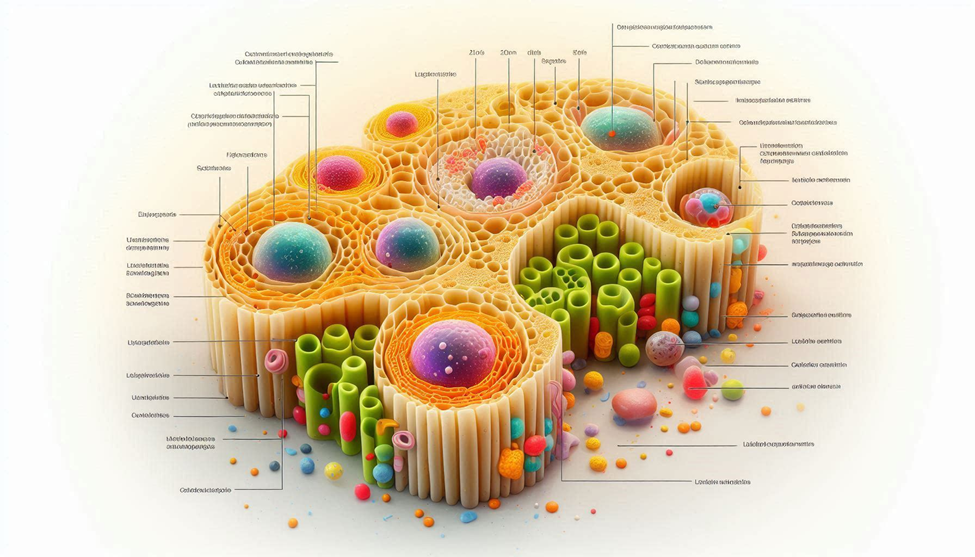
As research expands, lipid droplets are now implicated in diverse processes—from thermo genesis in brown adipose tissue to steroid hormone synthesis in endocrine tissues and viral replication in infected cells. Appreciating their central role in metabolic health provides a more precise, biology-driven understanding of weight regulation and energy balance.
This guide provides a comprehensive, highly detailed, deeply expanded overview of lipid droplet biology—its structure, formation, regulation, nutritional modulation, and metabolic significance—supported by advanced scientific analysis and professionally integrated concepts.
1.1 What Are Lipid Droplets?
Lipid droplets are intracellular organelles composed primarily of neutral lipids—triacylglycerols (TAGs) and cholesterol esters—surrounded by a unique phospholipids monolayer studded with regulatory proteins. Unlike other organelles with belayed membranes, LDs have a monolayer because their hydrophobic core requires no internal aqueous separation.
LDs vary dramatically in size, function, and behavior depending on the cell type:
- Adiposities may contain a single, massive LD that occupies up to 90% of the cell’s volume.
- Hepatocytes contain multiple LDs involved in lipoprotein assembly and lipid detoxification.
- Skeletal muscle LDs serve as rapid-mobilizing fuel during exercise.
- Immune cells use LDs for inflammatory mediator synthesis.
LDs are also dynamic: they can emerge rapidly in response to high-energy intake, shrink with biolysis, and undergo structural remodeling with stress or hormonal signaling. This adaptability makes them essential for metabolic resilience and cellular survival.
1.2 Why Lipid Droplet Biology Matters
The biology of LDs is essential for understanding obesity, metabolic syndrome, and energy physiology because:
- They determine how efficiently the body stores excess calories: Different individuals vary in LD expansion capacity; impaired expandability leads to fat spilling over into ectopic tissues such as the liver and muscle, driving insulin resistance.
- They control lipid flux between storage and oxidation: LDs coordinate with mitochondria and paroxysms to determine whether fat is burned or stored.
- They influence hormonal sensitivity: LD proteins modulate insulin signaling, glucocorticoid responses, catecholamine sensitivity, and adipokine secretion.
- They regulate inflammation: LDs house enzymes that generate inflammatory lipids (eicosanoids). The size and number of LDs change dramatically during immune activation.
- They integrate nutrient and environmental signals: LD behavior is shaped by feeding patterns, macronutrient composition, physical activity, thermal stress, sleep, and circadian rhythms.
Lipid droplets are therefore not merely storage depots—they are dynamic hubs governing metabolic health, cellular stress responses, and systemic energy balance.
1.3 The Cellular Architecture of a Lipid Droplet
LD structure is simple yet biochemically sophisticated:
Core (hydrophobic interior)
- Triacylglycerols (TAGs)
- Diacylglycerols (DAGs)
- Cholesterol esters
- Fatty acids
Monolayer membrane
A phospholipids monolayer, not a belayed, composed mainly of:
- Phosphatidylcholine
- Phosphatidylethanolamine
- Phosphatidic acid
Protein coat
This is what makes LDs metabolically interactive. Key proteins include:
- Peril pins (PLIN1-5) — master regulators of LD stability and biolysis
- ATGL (adipose triglyceride lipase) — initiates TAG breakdown
- HSL (hormone-sensitive lipase) — hydrolyzes DAG
- CGI-58 — essential co activator of ATGL
- Lipases, kinesis, transport proteins, and fusion machinery
These proteins control droplet size, biolytic sensitivity, and the capacity to store or release fat. The protein composition of LDs shifts dramatically depending on nutrient state and hormonal environment.
1.4 The Endoplasmic Reticulum: Birthplace of Lipid Droplets
Lipid droplets originate from the endoplasmic reticulum (ER), where enzymes synthesize triglycerides. Newly formed TAG molecules accumulate within the ER belayed until they form small neutral-lipid lenses.
When these lenses grow large enough, they bud off into the cytosol, forming nascent LDs.
Key steps:
- Neutral lipid synthesis
- TAGs and cholesterol esters are produced in the ER membrane.
- Lipid lens formation
- Neutral lipids accumulate between ER membrane leaflets.
- Monolayer formation
- ER phospholipids form the LD surface.
- Droplet budding and growth
- LDs detach into the cytosol or remain ER-connected for continued lipid exchange.
This ER-LD interaction remains dynamic even after budding, allowing for ongoing lipid trafficking, membrane expansion, and protein localization.
1.5 Growth and Expansion of Lipid Droplets
Once formed, LDs expand in three ways:
- Local TAG synthesis at the LD surface: LD-associated enzymes continue to build TAGs directly on the monolayer, enlarging the droplet.
- Fusion of multiple small droplets: Smaller LDs may merge into larger ones, especially in adiposities.
- Uptake of lipids from the ER or cytosolic sources: LDs can absorb lipids from lipoproteins or other organelles.
Peril pins regulate this expansion by shielding stored lipids and controlling access of lipases.
Expansion is adaptive: it prevents lip toxicity by storing potentially harmful fatty acids in safe, neutral forms.
1.6 Lipid Droplet Heterogeneity: Not All Droplets Are the Same
LDs vary based on:
1. Cell type
- White adiposities: massive storage droplets
- Brown adiposities: many small droplets for rapid mobilization
- Liver cells: multifunctional droplets linked to lipoprotein production
- Muscle cells: exercise-responsive droplets near mitochondria
2. Droplet size
- Large, unilocular droplets store long-term energy.
- Smaller, multilocular droplets support rapid utilization.
3. Protein composition
LD protein signatures differ under:
- Fasting
- High-fat feeding
- Exercise training
- Inflammation
- Insulin resistance
4. Metabolic function
Some LDs are primarily storage depots; others are oxidation-linked, inflammatory-linked, or endocrine-linked.
This heterogeneity allows tissues to meet diverse metabolic demands with precision.
2. Lip genesis: How Lipid Droplets Store Energy
Lip genesis is the process by which cells synthesize fatty acids and triglycerides for storage in lipid droplets. This process integrates signals from nutrient availability, hormonal cues, and energy demand. In adiposities, hepatocytes, and even skeletal muscle, lip genesis is tightly regulated to prevent lip toxicity while ensuring energy reserves for periods of scarcity.
2.1 De Novo Lip genesis
De novo lip genesis (DNL) converts excess carbohydrates into fatty acids. Glucose is metabolized into acetyl-Coal, which serves as the substrate for fatty acid syntheses (FAS) and acetyl-Coal carboxyl’s (ACC). The resulting palpitate can be elongated or desiderated before incorporation into triglycerides.
- Hormonal regulation: Insulin up regulates biogenic enzymes and promotes LD expansion, while glucagon suppresses lip genesis.
- Nutrient influence: High carbohydrate intake, particularly fructose, increases DNL in the liver, contributing to hepatic statuses if unchecked.
2.2 Etherification and TAG Formation
Fatty acids are esterifies to glycerol-3-phosphate by a series of enzymes including GPAT, AGPAT, and DGAT. Triglycerides are then packaged into lipid droplets. The efficiency of this process is critical: inadequate storage capacity can lead to free fatty acid accumulation, mitochondrial stress, and insulin resistance.
2.3 Role of Peril pins in Storage
Peril pins (PLIN1–5) coat LDs and regulate biolytic accessibility. PLIN1, primarily in white adiposities, protects stored triglycerides from basal lipase activity while permitting rapid biolysis upon adrenergic stimulation. PLIN2 and PLIN3 are ubiquitous and modulate droplet stability and growth.
2.4 Biolysis: Mobilizing Stored Energy
Biolysis is the breakdown of stored triglycerides into free fatty acids (FFAs) and glycerol for energy production. It is catalyzed primarily by:
- ATGL (adipose triglyceride lipase) – initiates TAG hydrolysis
- HSL (hormone-sensitive lipase) – hydrolyzes DAG to MAG
- MAG lipase – completes the breakdown to glycerol and FFAs
Biolysis is tightly regulated by hormones: catecholamines activate biolysis via camp/PKA signaling, while insulin inhibits it. Deregulation can lead to excessive FFA release, contributing to ectopic fat deposition and metabolic dysfunction.
2.5 Lipid Droplets and Mitochondrial Interaction
LDs are not isolated storage units; they communicate extensively with mitochondria. Physical contacts facilitate:
- Direct lipid transfer for β-oxidation
- Regulation of mitochondrial dynamics during energy demand
- Integration of redo signaling to prevent oxidative stress
Exercise, fasting, and adrenergic stimulation increase LD–mitochondria contacts, enhancing rapid mobilization of energy.
2.6 Lipid Droplet Proteins beyond Peril pins
Beyond peril pins, LD-associated proteins include:
- CIDE family proteins – regulate droplet fusion and apoptosis-linked lipid metabolism
- Fsp27/CIDEC – mediates unilocular LD formation in white adiposities
- Seeping – critical for droplet biogenesis and size regulation
- Rib proteins – control droplet trafficking and interactions with endoscopes
These proteins determine droplet size, mobility, biolysis sensitivity, and cross-talk with other organelles, illustrating LDs’ role as active metabolic hubs.
2.7 Lipid Droplet Dynamics in Different Tissues
- White adipose tissue (WAT): Long-term energy storage, endocrine signaling (lepton, adiponectin), metabolic buffer
- Brown adipose tissue (BAT): Multilocular LDs support thermo genesis; LD–mitochondria interaction is critical for heat production
- Liver (hepatocytes): LDs handle lipoprotein assembly, DNL, detoxification; excessive accumulation leads to non-alcoholic fatty liver disease (NAFLD)
- Skeletal muscle: LDs near mitochondria provide rapid energy for contraction; endurance training increases LD number and oxidative capacity
- Macrophages and immune cells: LDs serve as reservoirs for inflammatory mediators (eicosanoids) during immune activation
Tissue-specific differences reflect functional specialization, metabolic flexibility, and the integrated role of LDs in systemic energy balance.
3. Hormonal Regulation of Lipid Droplets
Lipid droplet metabolism is profoundly influenced by endocrine signals. Hormones dictate whether triglycerides are stored or mobilized, allowing tissues to adapt to nutritional and energetic status.
3.1 Insulin: Master Storage Hormone
Insulin is the primary anabolic signal promoting LD expansion. It stimulates:
- Glucose uptake via GLUT4 transporters in adiposities and muscle
- Lip genesis through activation of ACC and FAS
- Inhibition of biolysis by dephosphorylating HSL and suppressing ATGL activity
Chronic hyperinsulinemia can lead to excessive LD accumulation, particularly in liver and visceral adipose tissue, contributing to insulin resistance.
3.2 Catecholamine’s: Mobilizing Energy Stores
Epinephrine and nor epinephrine activate β-adrenergic receptors, increasing camp and activating PKA. This cascade phosphorylates peril pins and HSL, promoting biolysis. LDs in BAT are especially responsive, providing fatty acids for mitochondrial oxidation and thermo genesis.
3.3 Glucocorticoids: Dual-Role Hormones
Cortical modulates both storage and mobilization:
- Enhances lip genesis and LD expansion in visceral adipose tissue
- Stimulates biolysis during fasting and stress
- Excess cortical contributes to central obesity and metabolic deregulation
3.4 Growth Hormone and Other Peptides
Growth hormone (GH) and natriuretic peptides promote biolysis and enhance fatty acid availability for energy. Adipokines such as lepton and adiponectin modulate LD metabolism indirectly, regulating systemic energy balance and insulin sensitivity.
3.5 Nutrient and Dietary Influences on LD Dynamics
Dietary composition profoundly affects LD formation and function:
- High-carbohydrate diets stimulate DNL and liver LD expansion
- High-fat diets increase adiposity LD size and number
- Omega-3 fatty acids promote smaller, metabolically active LDs and improve mitochondrial oxidation
- Caloric restriction reduces LD size, increases biolytic responsiveness, and enhances metabolic flexibility
Meal timing and frequency also modulate LD dynamics via circadian hormone patterns.
3.6 Lipid Droplets in Metabolic Disease
Deregulation of LD biology underpins multiple metabolic disorders:
- Obesity: LD expansion exceeds safe storage capacity, leading to ectopic lipid deposition
- NAFLD: Excess hepatic LDs impair insulin signaling and increase oxidative stress
- Type 2 diabetes: Impaired LD turnover in muscle reduces oxidative flexibility
- Atherosclerosis: Foam cells are macrophages overloaded with LDs, promoting plaque formation
Targeting LD regulation is emerging as a promising therapeutic avenue for metabolic diseases.
3.7 Clinical Implications and Nutritional Strategies
Understanding LD biology informs practical interventions:
- Exercise: Enhances LD–mitochondria contacts in muscle, supporting efficient fatty acid oxidation
- Macronutrient balance: Adequate protein and fiber stabilize blood glucose, reducing hepatic DNL
- Sleep optimization: Prevents cortical-induced LD expansion in visceral fat
- Targeted nutrient supplementation: Omega-3s, polyphones, and medium-chain triglycerides modulate LD size and biolysis efficiency
Integrating these strategies aligns lifestyle practices with biological mechanisms, supporting metabolic health beyond calorie restriction or willpower.
4. Lipid Droplets and Organelle Interactions
Lipid droplets are not isolated structures; they form dynamic physical and functional interactions with other organelles, integrating cellular metabolism and stress responses.
4.1 LD–Mitochondria Contacts
- Facilitate rapid fatty acid transfer for β-oxidation
- Promote energy production during exercise, fasting, or cold exposure
- Regulate mitochondrial dynamics and prevent oxidative stress via localized lipid buffering
4.2 LD–Endoplasmic Reticulum Communication
- ER remains the site of LD biogenesis and lipid exchange
- ER–LD contact sites allow triglyceride trafficking, phospholipids remodeling, and expansion of LD monolayer’s
- Dysfunction can lead to ER stress, lip toxicity, and impaired protein folding
4.3 LD–Peroxisome Interactions
- Paroxysms metabolize very-long-chain fatty acids stored in LDs
- LD–peroxisome contacts enhance lipid catabolism and reduce reactive oxygen species
4.4 LD–Lissome Crosstalk
- LDs can be degraded through lipophagy, a form of selective autophagy
- Liposomal degradation releases fatty acids for energy production during fasting or nutrient stress
- Lipophagy deregulation contributes to hepatic statuses and impaired metabolic flexibility
4.5 Lipid Droplets in Inflammation and Immune Function
LDs are central hubs for inflammatory signaling:
- Store precursors for eicosanoids and prostaglandins
- Regulate cytokine production in macrophages and neutrophils
- Increase in number and size during infection or metabolic stress
- Serve as platforms for host-pathogen interactions (e.g., viral replication)
LDs thus bridge metabolism and immunity, highlighting their dual role in health and disease.
4.6 Stress, Oxidative Damage, and LD Adaptation
Cells use LDs to buffer stress:
- Sequester excess fatty acids to prevent lip toxicity
- Protect membranes from oxidative damage
- Coordinate stress responses with mitochondrial and ER signaling
Chronic stress, nutrient excess or oxidative load can overwhelm LD capacity, contributing to metabolic disorders.
4.7 Advanced Clinical and Metabolic Applications
Understanding LD dynamics opens therapeutic and lifestyle applications:
- Metabolic flexibility training: Exercise and nutrient timing optimize LD utilization for energy
- Targeted pharmacology: Modulators of ATGL, peril pins, and lipophagy pathways are under investigation for obesity and NAFLD
- Nutritional modulation: Balanced macronutrient intake, omega-3s, polyphones, and medium-chain fats influence LD size and biolytic responsiveness
- Precision medicine: Identifying individual LD profiles in adipose and liver tissue may predict metabolic risk and personalize interventions
LD biology offers actionable insights that go far beyond simple calorie restriction, aligning interventions with cellular physiology for sustainable health outcomes.
5. Lipid Droplet Turnover and Metabolic Flexibility
The balance between lipid droplet formation and breakdown underlies metabolic flexibility—the ability of cells and tissues to adapt fuel utilization to changing energy demands. Rapidly mobilizable LDs in muscle and brown adipose tissue support acute energy needs, while long-term storage LDs in white adipose tissue buffer excess calories. Turnover is regulated by the coordinated action of lipases, peril pins, autophagy, and organelle crosstalk, ensuring that energy supply matches demand without triggering lip toxicity.
5.1 Nutritional Modulation of Lipid Droplets
Diet profoundly influences LD biology:
- High-carbohydrate diets: Promote de novo lip genesis, increasing hepatic LDs
- High-fat diets: Stimulate adiposity LD expansion and alter peril pin expression
- Protein intake: Enhances satiety and modulates insulin signaling, indirectly regulating LD growth
- Polyunsaturated fats: Reduce droplet size, enhance mitochondrial oxidation, and improve metabolic efficiency
- Caloric restriction: Reduces LD size and enhances biolytic responsiveness, promoting longevity and metabolic health
Meal timing and circadian alignment also influence LD turnover, linking metabolic rhythm to cellular lipid handling.
5.2 Exercise and Lipid Droplet Dynamics
Physical activity remodels LDs across tissues:
- Skeletal muscle: Endurance training increases the number of small, mitochondria-associated LDs, enhancing fatty acid oxidation
- Brown adipose tissue: Exercise-induced adrenergic signaling increases LD–mitochondria contacts, boosting thermo genesis
- Adipose tissue: Exercise promotes smaller, more metabolically active LDs, improving insulin sensitivity
LD remodeling via exercise illustrates how behavioral interventions can synergize with cellular metabolism.
5.3 Aging, Lipid Droplets, and Metabolic Decline
Aging affects LD number, size, and turnover:
- White adiposities enlarge, reducing expandability and increasing ectopic lipid deposition
- Skeletal muscle LDs become less mitochondria-associated, reducing metabolic flexibility
- Hepatic LDs accumulate, contributing to NAFLD and insulin resistance
- Impaired lipophagy decreases efficient lipid mobilization
Nutritional strategies, exercise, and targeted interventions can mitigate age-related LD deregulation.
5.4 Lipid Droplets in Systemic Energy Homeostasis
LDs influence whole-body energy balance:
- Adipose tissue: Stores triglycerides, secretes adipokines, buffers postprandial lipids
- Liver: Regulates circulating lipid availability and lipoprotein assembly
- Muscle: Provides rapid fuel for contraction and maintains oxidative capacity
- Immune system: Coordinates energy supply during inflammation and stress
Disruption in any tissue’s LD function can trigger systemic metabolic dysfunction, demonstrating their centrality in physiology.
5.5 Pharmacological and Molecular Targets
Key LD-associated proteins and pathways are being explored therapeutically:
- ATGL activators/inhibitors to modulate biolysis
- Peril pin modulators to regulate droplet accessibility
- Lipophagy enhancers to improve turnover and prevent ectopic fat accumulation
- Fsp27/CIDEC inhibitors for adiposity droplet remodeling
Precision targeting of LD biology holds potential for obesity, diabetes, and NAFLD treatments.
5.6 Integrating Lipid Droplet Knowledge into Lifestyle Design
Practical interventions grounded in LD biology include:
- Balanced macronutrients to stabilize insulin and regulate LD expansion
- Nutrient timing to align lip genesis and biolysis with circadian rhythms
- Regular exercise to remodel LDs and enhance mitochondrial association
- Sleep and stress management to modulate cortical-driven LD expansion
- Supplementation with omega-3s, polyphones, and medium-chain triglycerides to optimize droplet size and turnover
This systems-based approach enables sustainable metabolic health by leveraging cellular physiology.
5.7 Future Directions in Lipid Droplet Research
Emerging areas include:
- Single-cell LD profiling: Identifying tissue-specific LD heterogeneity and metabolic risk
- Organelle interactomics: Mapping LD contacts with mitochondria, ER, paroxysms, and lissome
- Gene editing and synthetic biology: Modifying LD proteins to improve metabolic flexibility
- Nutraceutical development: Targeting LD pathways with dietary compounds
- Clinical imaging: Using advanced imaging to quantify LD dynamics in vivo
These directions promise to translate fundamental LD biology into precision interventions for metabolic disease prevention and treatment.
Conclusion
Lipid droplets, once considered passive storage depots, are now recognized as dynamic, multifunctional organelles central to energy homeostasis, cellular signaling, and systemic metabolic regulation. Their formation, growth, and breakdown are orchestrated by intricate networks of enzymes, structural proteins, and organelle interactions, all finely tuned by hormonal, nutritional, and environmental cues. The heterogeneity of LDs across tissues—from unilocular white adiposities to mitochondria-associated droplets in skeletal muscle and multilocular droplets in brown adipose tissue—reflects their specialized roles in storage, energy mobilization, thermo genesis, and immune function.
Understanding the biology of lipid droplets reframes metabolic health. Rather than relying on simplistic models of calorie balance or willpower, LD dynamics reveal that energy storage and release are tightly regulated physiological processes, shaped by neuroendocrine signals, macronutrient composition, and lifestyle factors such as exercise, sleep, and circadian alignment. Deregulation of LD turnover or expansion contributes to obesity, insulin resistance, non-alcoholic fatty liver disease, and cardiovascular risk, highlighting their clinical significance.
From a practical perspective, leveraging LD biology offers actionable strategies for metabolic optimization. Targeted dietary patterns, timed feeding, structured exercise, stress management, and specific nutrient interventions can enhance LD turnover, improve mitochondrial interactions, and promote metabolic flexibility. Moreover, ongoing research into LD-associated proteins, lipophagy pathways, and organelle communication holds promise for therapeutic innovation in metabolic disease.
Ultimately, lipid droplets exemplify how a deep understanding of cellular physiology can transform approaches to health. By integrating insights from molecular biology, endocrinology, and nutrition science, it is possible to design interventions that work in harmony with the body’s natural energy-regulatory mechanisms, enabling sustainable metabolic health, resilience, and longevity.
SOURCES
Fares, R.V. & Walther, T.C. (2009). Lipid droplets finally get a little respect. Cell, 139(5), 855–860.
Walther, T.C. & Fares, R.V. (2012). Lipid droplets and cellular lipid metabolism. Annual Review of Biochemistry, 81, 687–714.
Poll, A., Gross, S.P. & Parton, R.G. (2014). Biogenesis of the multifunctional lipid droplet: Lipids, proteins, and sites. Nature Reviews Molecular Cell Biology, 15(8), 507–521.
Bickel, P.E., Tansy, J.T. & Welted, M.A. (2009). Lipid droplets, adipose triglyceride lipase, and the biolytic barrier. Journal of Lipid Research, 50(Supple), S86–S91.
Lichtenberger, L.L. & Brown, D.A. (2007). Lipid droplets and cellular lipid homeostasis. Current Opinion in Cell Biology, 19(4), 363–368.
Cohen, P., Miyazaki, M., Scio, R.E., et al. (2008). Role of peril pin in lipid metabolism. Cell Metabolism, 8(6), 467–479.
Thai, A.R., Fares, R.V. & Walther, T.C. (2013). The biophysics and cell biology of lipid droplets. Nature Reviews Molecular Cell Biology, 14(12), 775–786.
Greenberg, A.S., Egan, J.J., Week, R.C., et al. (2001). Peril pin, lipid droplet proteins, and regulation of biolysis. Journal of Biological Chemistry, 276(7), 5201–5208.
Kimmel, A.R. & Sztalryd, C. (2016). The peril pins: Major cytosolic lipid droplet-associated proteins. Current Opinion in Cell Biology, 41, 109–116.
Sztalryd, C. & Brasaemle, D.L. (2017). Role of lipid droplet proteins in biolysis and signaling. Trends in Endocrinology & Metabolism, 28(6), 449–460.
Lichtenberger, L.L., Or, D.S. & Schaffer, J.E. (2001). Palpitate-induced lip toxicity: Mechanisms and protection by lipid droplets. Journal of Biological Chemistry, 276(42), 38084–38091.
Ago, Q., Fu, X., CSU, M., et al. (2017). Lipid droplet–mitochondria interactions in metabolism. Trends in Cell Biology, 27(6), 434–446.
Fechner, R., Made, F. & Katy, D. (2017). Cytosolic biolysis and lipid droplet dynamics. Nature Reviews Molecular Cell Biology, 18(12), 671–684.
Greenberg, A.S., Egan, J.J., Chang, M.C., et al. (1991). Lipase activation and lipid droplet protein regulation. Journal of Lipid Research, 32(12), 1955–1966.
Martin, S., Parton, R.G. (2006). Lipid droplets: A unified view of a dynamic organelle. Nature Reviews Molecular Cell Biology, 7(5), 373–378.
Kramer, N., Fares, R.V. & Walther, T.C. (2013). Balancing the fat: Lipid droplets and metabolic homeostasis. Trends in Cell Biology, 23(5), 251–259.
Boltzmann, J.A. & Carvalho, P. (2019). Dynamics and functions of lipid droplets. Nature Reviews Molecular Cell Biology, 20(3), 137–155.
Cain, Y., Wang, Y., Zing, J., et al. (2020). Lipid droplet turnover in health and disease. Cell & Molecular Life Sciences, 77(16), 3193–3212.
Herder, E., Tot, M. (2012). Emerging roles of lipid droplets in cell biology. Cellular Microbiology, 14(9), 1419–1428.
Nguyen, T.B., Boltzmann, J.A., et al. (2017). Lipid droplets as multifunctional organelles. Annual Review of Cell and Developmental Biology, 33, 491–510.
Herzog, R., et al. (2016). Proteomics of lipid droplets in different tissues. Molecular & Cellular Proteomics, 15(2), 549–561.
Meyers, M.P., et al. (2021). Lipid droplet interactions with mitochondria: Implications for metabolism. Trends in Endocrinology & Metabolism, 32(2), 123–136.
Kasson, A., et al. (2013). CIDE proteins and lipid droplet fusion in adiposities. Journal of Lipid Research, 54(2), 315–322.
Bistro, P., et al. (2012). SNARE-mediated interactions between LDs and organelles. Cell Metabolism, 16(4), 394–403.
Lichtenberger, L.L., et al. (2007). Lipid droplets as metabolic and stress-response organelles. Journal of Lipid Research, 48(11), 2399–2410.
HISTORY
Current Version
Nov 25, 2025
Written By
ASIFA

0 Comments